
Temperature Dependent Sex Determination
I recently ran into a few scenarios where my broods produced highly skewed sex ratios. For the most part, these anomalies were heavily biased toward males. Recently, however, I had a couple of broods that resulted in 100% females. This was incredibly puzzling, which prompted me to dig into the potential impact of temperature on the sex of guppy fry. I was initially very skeptical of Temperature-Dependent Sex Determination (TDSD) in guppies, but I've come to realize it may hold true under specific conditions.
To explore this, I wanted to provide a synopsis of a commonly referenced 2006 study titled, Effect of temperature on sex ratio in guppy Poecilia reticulata (Peters 1860) by Ismihan Karayücel, Orhan Ak, and Sedat Karayücel.
Note: None of the research presented on this page is mine. If you would like to read the original paper, you can find it here. I am simply repackaging their findings through the lens of what might have occurred in my own all-female broods. You can find the companion video for the discussion below here.
Embryos vs. Newborn Fry: When is Temperature Most Potent?
If temperature influences the final sex ratio of a brood, a natural follow-up question is: When is the window of susceptibility? Are guppies more vulnerable to thermal shifts as developing embryos in the womb, or as newborn fry? The authors tested both stages using two distinct experiments:
Experiment 1 (Embryo Susceptibility): Pregnant females were subjected to temperature treatments during the latter half of their gestation.
Experiment 2 (Fry Susceptibility): Newborn fry were subjected to temperature treatments for the first 11 days of life.
Unfortunately, during the embryo testing, many of the pregnant females died shortly after being placed in the experimental tanks. Because pregnant females are highly sensitive to external environmental stressors, this first experiment was largely inconclusive. Consequently, this synopsis will focus primarily on the findings from the newborn fry experiments.
Establishing a "Labile" Baseline
The authors were highly systematic in their approach. Before diving into thermal testing, they needed to ensure their guppy strain was actually susceptible to temperature shifts. Interestingly, certain guppy strains are completely immune to temperature-induced sex changes. The authors referred to temperature-susceptible guppies as labile. Once they established a labile baseline group, they set up the experiment using 18 adult females. They chose to test six different target temperatures using three females per treatment group, alongside a control group maintained at 26°C (78.8°F):
Cooler Treatments: 19°C (66.2°F), 21°C (69.8°F), and 22.5°C (72.5°F)
Warmer Treatments: 29°C (84.2°F), 32°C (89.6°F), and 35°C (95°F)
To establish a baseline control, the first brood from each of the 18 females was raised strictly at the 26°C (78.8°F) control temperature. The subsequent broods from these identical females were used as the experimental groups. These fry were placed into temperature-controlled containers for the first 11 days of life before being returned to the 26°C (78.8°F) control environment for the remainder of the 50-day experiment.
Analyzing Fry Survival Rates
Before looking at sex ratios, the authors recorded fry survival. Table 2 (reproduced below) outlines the original survival totals. The first set of columns documents the survival rates of the control broods at day 11 and day 50. The subsequent columns show the survival rates of the heat- and cold-treated broods. To visualize these trends more clearly, I averaged the data points and calculated the standard deviations for all 18 control replicates, as well as the triplicates for each of the six temperature treatments (Graph Below).
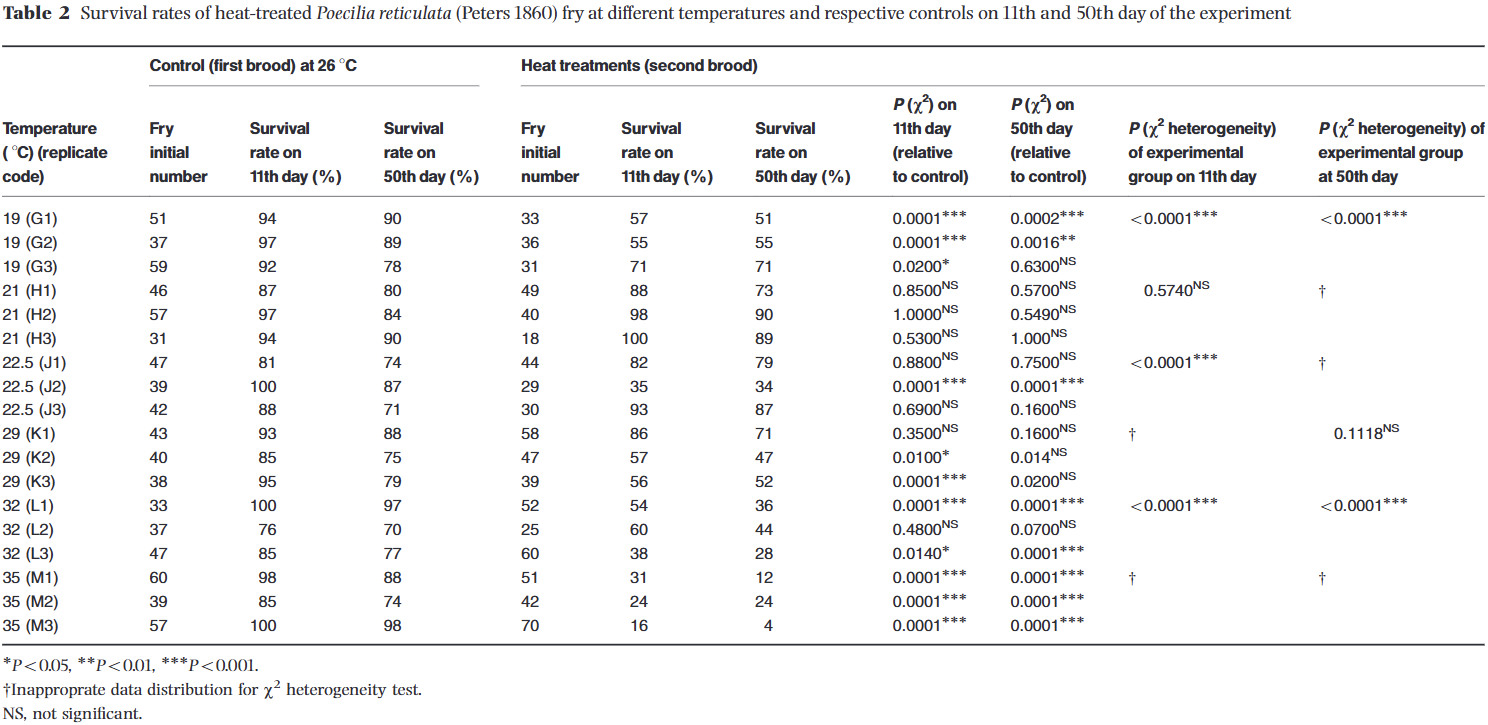
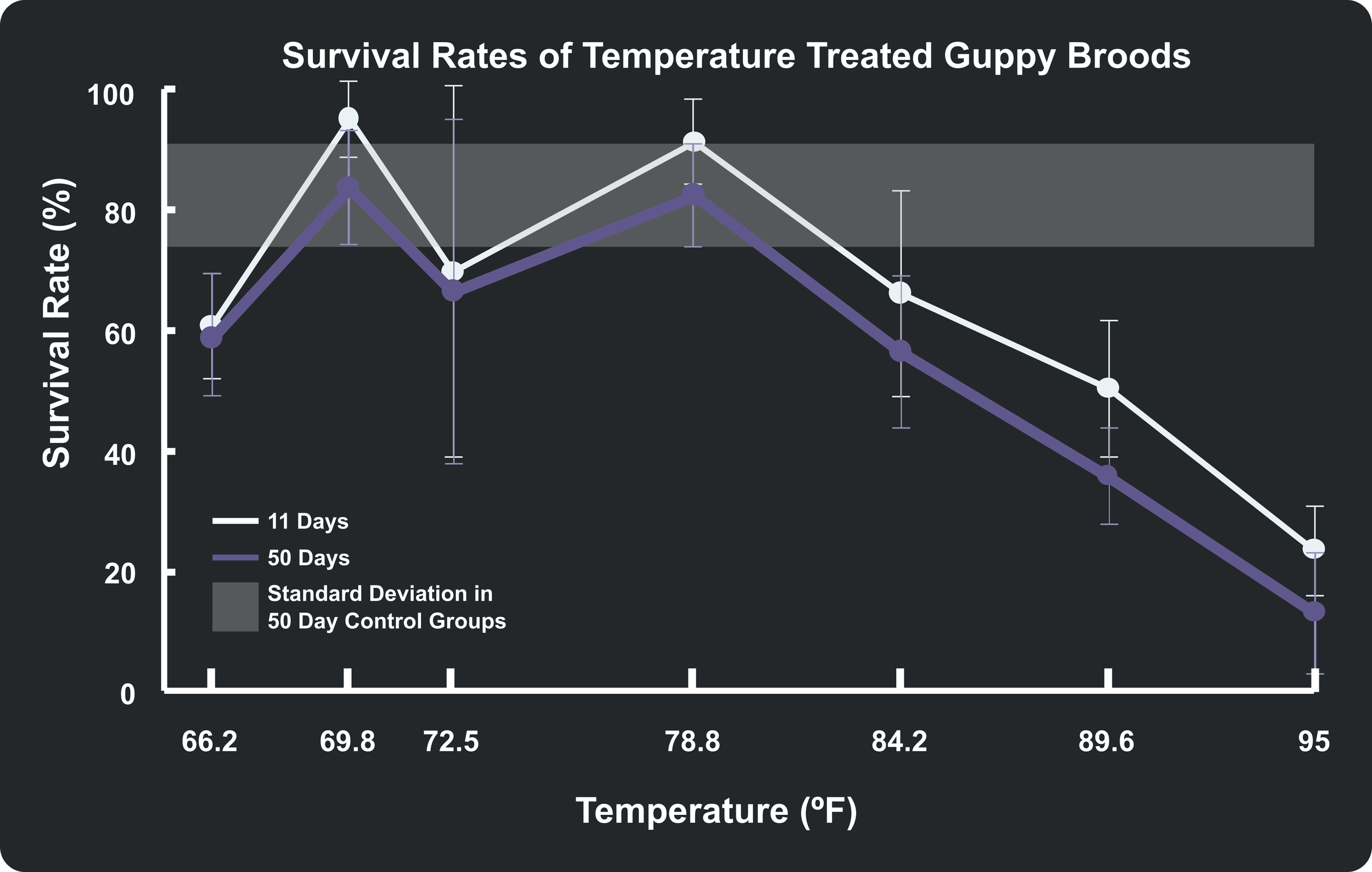
Unsurprisingly, the chart demonstrates that fry survival decreases as temperatures move further away from the natural baseline. To provide a clear visual benchmark, I added a shaded gray bar representing the standard deviation region of the 18 control groups; any data points lying outside of this band suggest a meaningful departure from normal survival expectations.
While overall mortality naturally increased between day 11 and day 50, the core survival trends across the temperature spectrum remained remarkably consistent. A notable feature of the graph is the wide error bars at 22.5°C (72.5°F). This high variance is driven entirely by replicate group J2 (Table 2), which suffered a much lower survival rate compared to the J1 and J3 groups at that same temperature.
While the authors utilized more rigorous chi-square χ2 statistical analyses to determine true significance compared to my simple averages and standard deviations, my visual model aligns closely with their findings. The data points that the authors flagged as highly significant (marked with asterisks in Table 2) at day 50 match up beautifully with the trends outside the shaded baseline on my graph. The only discrepancy occurs in the 29°C (84.2°F) treatment (Group K); my plot places these points slightly outside the normal control region, whereas the authors' statistical analysis determined the variation at 50 days to be non-significant (NS).
Moving from Survival to Sex Ratios
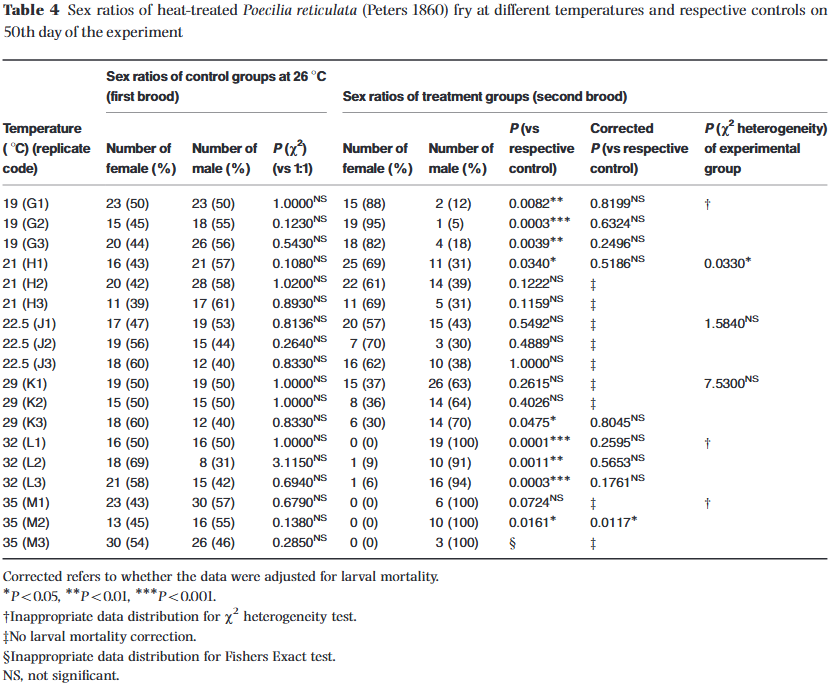
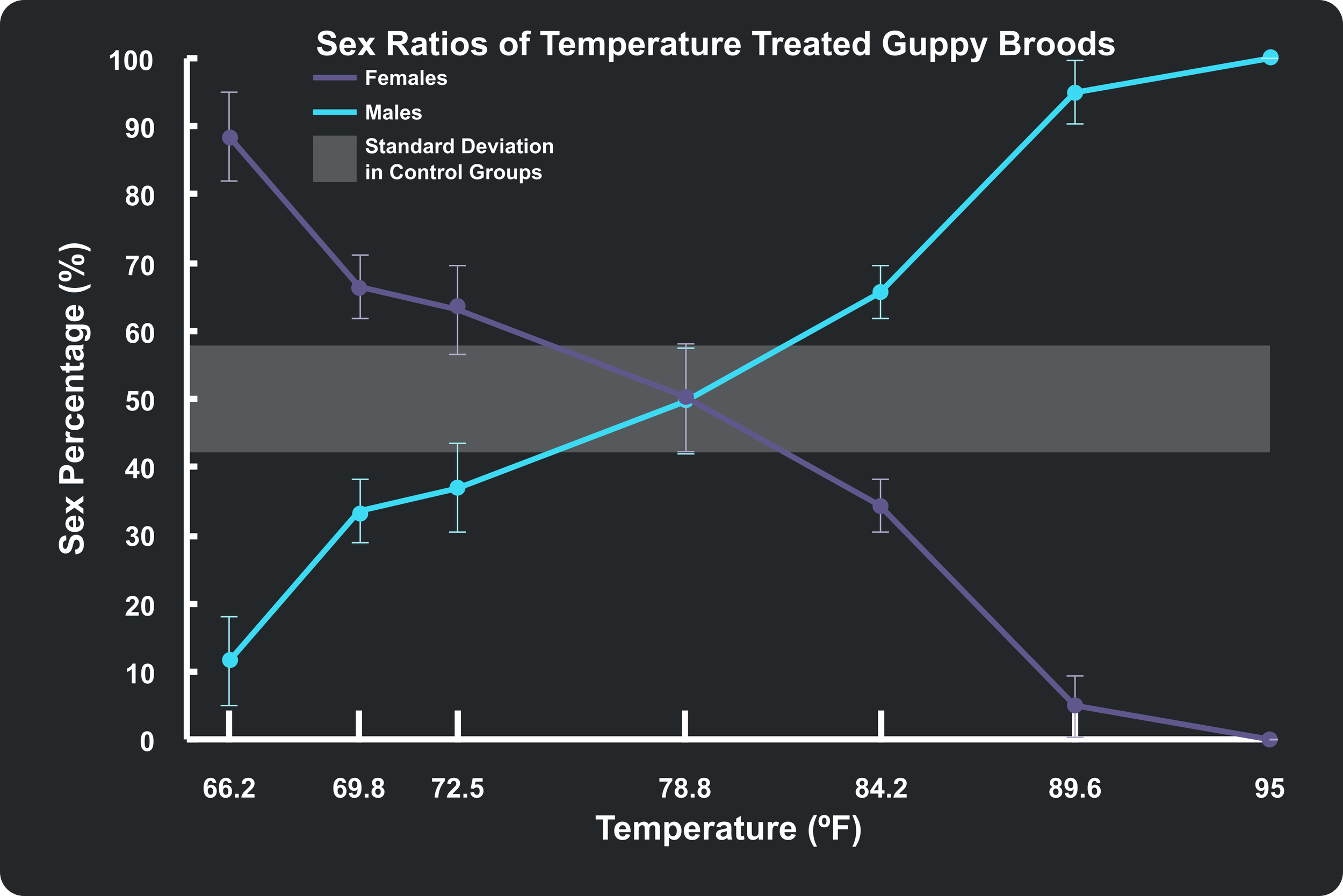
Once the survival rates were accounted for, the next major objective was analyzing how these temperature treatments shifted the actual sex ratios of the surviving fish. The researchers reported their findings in Table 4 (reproduced below) by showing the percentage of males and females within each brood. Again, I went ahead and collected the averages and standard deviations of this data and put them together into the plot below. I also added a shaded region for the standard deviation of the control group, which is centered around 50%. Data falling within this 50% region tells us that the sex ratios for those data points were largely unaffected by temperature. Looking at this plot, we can clearly see that the male and female ratios of each brood are definitely affected by temperature on both the cold and hot ends of the spectrum. The plot shows that there were more females when the temperatures were below 21°C (69.8°F) and more males when the temperatures were above 29°C (84.2°F). Note that because these ratios must add up to 100%, the lines representing the male and female ratios on my plot are effectively mirror images of each other.
This finding is quite astonishing and very drastic. Going back to my own brood from Cross 22 that resulted in 100% females, the data suggests that a possible explanation for this anomaly was temperature-driven. And it very well might have been—but more on that later.
To determine whether a skewed ratio was a true environmental anomaly or just a product of random chance, the authors ran the data through a strict statistical hierarchy based on individual brood sizes. They used standard Chi-squared tests for large broods, Yates-corrected tests for medium broods, and Fisher’s Exact tests for tiny groups.
I wanted to make sure I understood their data and ensure my simpler averages and standard deviations weren't telling a different story. So, I sat down to independently replicate their tables in a spreadsheet. However, I noticed some fascinating mathematical quirks and formatting errors hidden inside their published data. While these typos don't change the study's high-level biological conclusions, tracking them down felt like a true forensic investigation. If you love statistical formulas and want to see the exact data entry mistakes the authors made to get their published numbers, check out the companion piece: TDSD - Statistical Audit. Otherwise, let's skip the formulas and look at what this means for the biology of the fish.
The "Silent Killer" Hypothesis: Larval Mortality Correction
With the math validated, we can address the core biological mystery: Are temperature extremes actively flipping the sex of the fish, or are they simply killing off one sex more effectively than the other? To isolate environmental sex selection from sex-dependent mortality, researchers utilized a method called a Larval Mortality Correction. Think of this as a mathematical stress test for data. For any brood displaying a heavily skewed sex ratio, every single fry that died during the 50-day window is "revived" on paper and assigned to whichever sex was in the minority. If a skewed ratio was merely caused by temperature killing off a more fragile sex, this extreme overcorrection will completely flatten the variance and pull the graph right back into a balanced, standard 50/50 distribution (represented by the shaded region on my graph or a statistically non-significant value in the authors' Table 4).
When this overcorrection is applied to the colder temperature treatments, that is exactly what happens. If we look at the Larval Mortality Corrected plot below, the data points shift directly back into the expected 50/50 baseline regime. This strongly implies that in cold environments, the skewed ratios are driven by Temperature-Dependent Sex Mortality—males are simply less resilient to cold water as newborns.
However, the high-temperature treatments tell a vastly different story. Even after applying the maximum possible larval mortality correction—giving every deceased fry the benefit of the doubt and counting them as female—the final proportions actually flipped to suggest there should have been more females than males. This is not a realistic biological expectation, but it tells us that temperature alone cannot account for the heavy male-biased ratios that were originally reported.
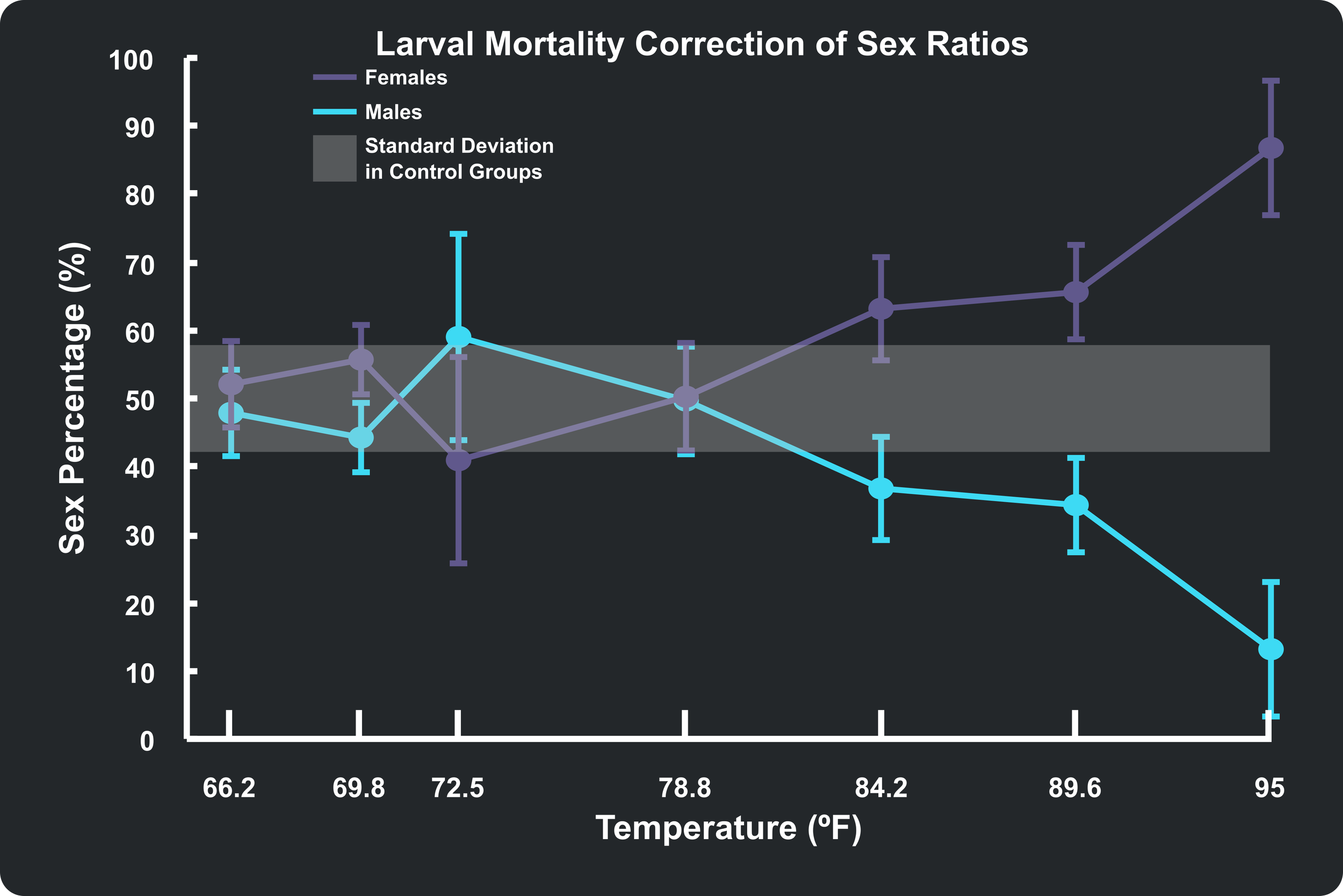
The data in my plots align closely with the values the authors reported in their Table 4, but not exactly. The authors reported that the only temperature treatment that resulted in non-significant values after correction was 35°C (95°F). Mine, which included an additional treatment group at 32°C (89.6°F), also sat outside the expected 50/50 region. To try and further refine my plots of the authors' data, I applied a Modified Correction to my own models. Knowing that the control groups suffered a baseline natural mortality rate of 17.3% even under ideal conditions (as shown in the Survival Rate plot above), I assigned that baseline percentage a standard 50/50 sex split and only applied the minority mortality correction to the remaining excess deaths.
You can see this additional correction applied to my plot below. It did indeed pull the 32°C (89.6°F) treatment group down into the 50/50 regime, matching the authors' Table 4 data. However, the lower temperature treatment groups got shifted further away from the 50/50 region. Even so, the error bars remain within the region and match very closely with the data reported by the authors. Overall, the bulk of the data is flattened out into the 50/50 shaded region, suggesting that temperature was indeed responsible for the sex bias (and mortality) of the young brood.
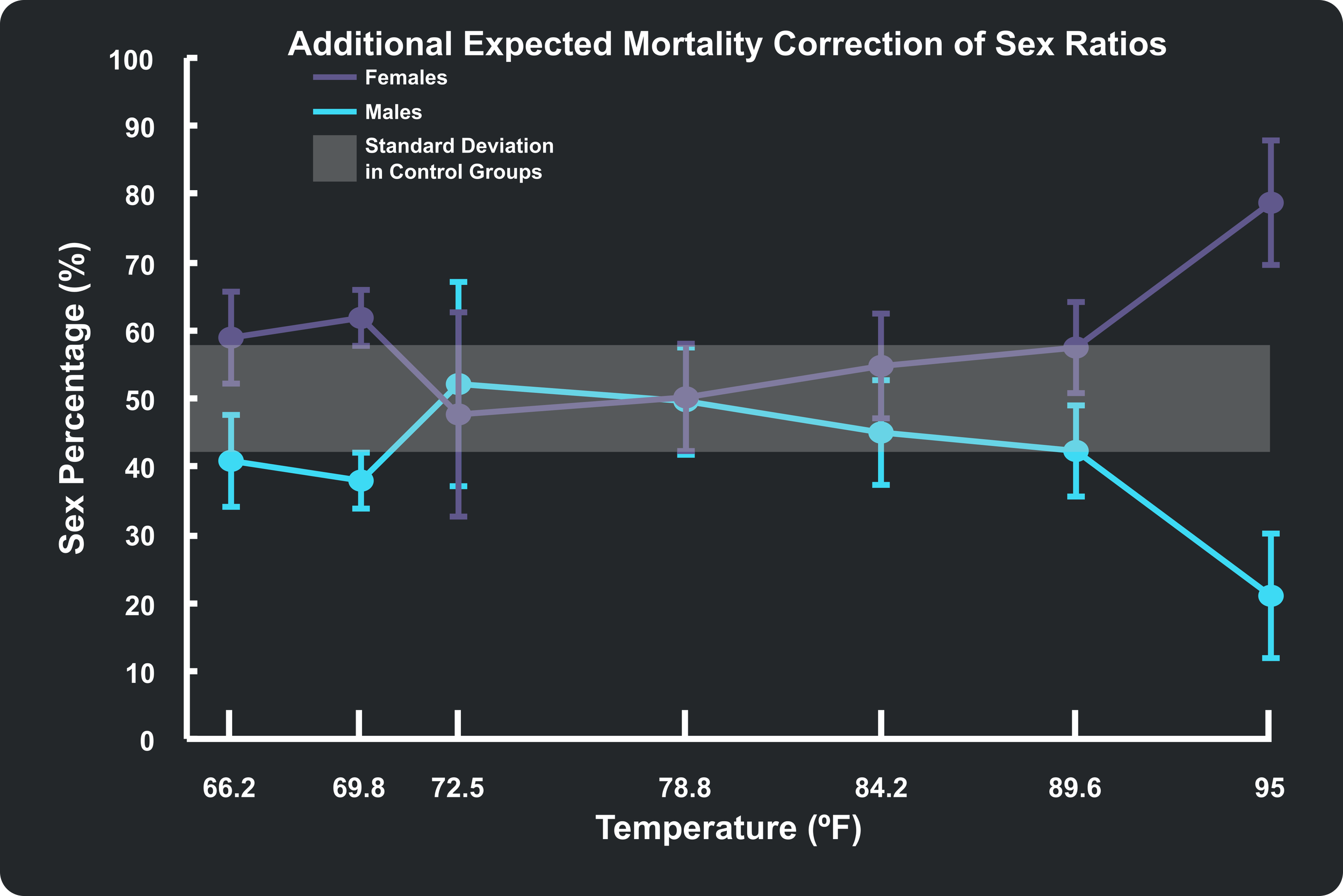
So what’s the verdict? Most of the sex-skewing observed in guppies across these temperatures is a product of sex-dependent mortality. But at the extreme upper thermal limit, there is a very real, statistically significant indication that something more complex—like true environmental sex reversal—is taking place.
The Genetic Anomaly: Are There "Sex-Reversed" Males?
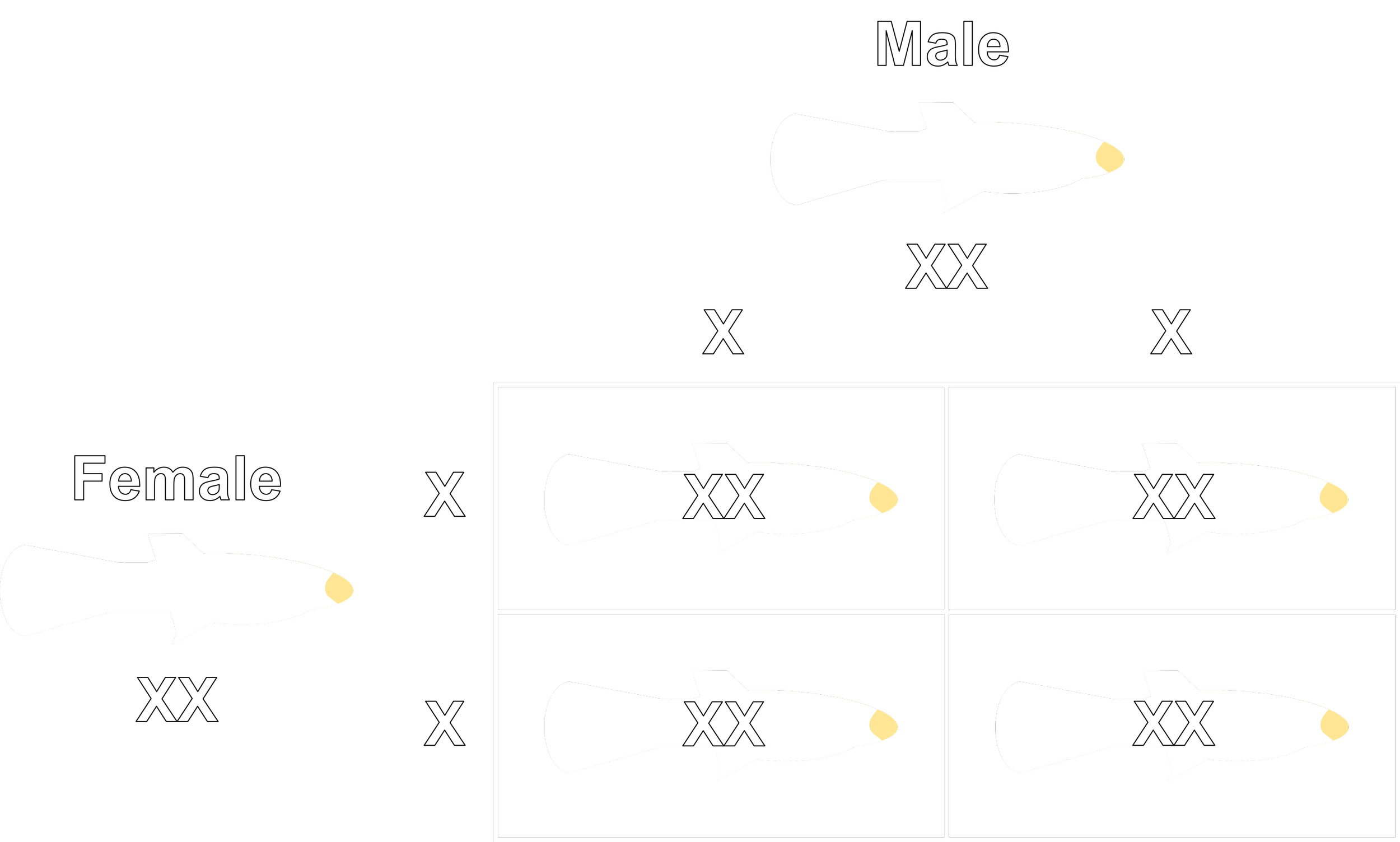
If extreme heat can actively override genetics and alter sex differentiation, it introduces an incredible phenomenon: the genotypic XX male. Normally, male guppies possess an X and a Y chromosome (XY), while females possess two X chromosomes (XX). If a high-temperature environment forces an XX embryo or fry to develop functional male anatomy, that fish becomes a phenotypic male while remaining genetically female.
This can easily be tested. If you breed a suspected, sex-reversed XX male back to a standard XX female, the Y chromosome is completely absent from the equation. Neither parent has a Y chromosome to pass down. Therefore, 100% of the resulting offspring must inherit an XX pairing and be born female (see the Punnett square below). The authors actually conducted progeny testing on the survivors of their high-temperature groups to look for this exact smoking gun. Interestingly, their specific test crosses did not yield any all-female anomalies, leading them to favor the mortality explanation over widespread sex determination changes.
However, finding a sex-reversed individual via random selection is incredibly difficult without a massive sample size or a visible, sex-linked physical trait to serve as a marker. It requires a great deal of luck, meaning the door to true temperature-dependent sex determination in specific sensitive lines isn't completely shut.
Back to Cross 22
This brings our scientific investigation full circle right back to my fish room and the "impossible" 100% female result of Cross 22. With the insights from the Karayücel paper, the answer might be hidden in the physical infrastructure of my tanks.
The grow-out tank for Cross 22 is located in my basement. Because the basement receives minimal heat from the household furnace, ambient room temperatures routinely drop below 65°F during the winter months. To combat this, I heat the water centrally in a large sump, which then pumps warm water into the individual project tanks via an inlet line. However, fluid dynamics and basic physics present a major challenge here: warm water rises, and cold water sinks. Because the overflow drain for this specific tank is positioned near the top at the back, the freshly warmed water from the sump was staying concentrated in the upper layer of the water column. When I tested the thermal layers with a digital thermometer during a cold spell, the top layer read a comfortable 76°F. The bottom layer of the tank, however, was sitting closer to a chilly 65°F.
Cross 22 was born on December 25th. Consequently, these newborn fry were swimming in water that was largely less than 70°F and probably closer to 65°F during their critical 11-day window of environmental susceptibility. Based on the definitive trends highlighted in the research paper, this stratified cold zone triggered extreme, sex-dependent male mortality. The males simply could not survive the localized thermal drop away from the surface of the tank, leaving behind a resilient, 100% female brood. While it is technically possible that the father himself was a rare product of high-temperature sex reversal from a hot summer brood—which would also yield 100% females—the cold basement gradient is a far more likely suspect.
Moving forward, the fix for this issue is straightforward and practical: adding dedicated aeration or sponge filters to actively circulate the water column, break up thermal stratification, and maintain a uniform temperature from top to bottom.
Overall, this was a fascinating deep dive. However, I do want to stress this again: not all guppy strains are susceptible to temperature. Some strains are completely immune to this, which the authors establish at the start of their article. This is likely due to a genetic resistance to temperature susceptibility, which the authors touch on toward the end of their paper.
Now that we’ve laid down this groundwork, we can begin to move toward talking about my 100% female brood from Cross 26, which can't be explained entirely by this temperature phenomenon…